Discussion
Unfortunately, Gyroporus has been the locus for some particularly sloppy scientific publications over the last 60 years or so. I should say right away that I mean no personal offense to any of the mycologists responsible for these publications, and that I have made many errors in my publications over the years. Also, many of the authors of the papers I'm about to criticize have done excellent work in other publications. But I feel it's important to document what the Gyroporus problems are, so that corrections can be made.
North American Species in the Gyroporus castaneus Complex
In 2020 Davoodian et al. described two new North American species in the Gyroporus castaneus complex: Gyroporus smithii and Gyroporus borealis; additionally the authors provided what they viewed as further support for Gyroporus purpurinus, which two of the authors had described earlier (Davoodian & Halling 2013). The 2020 paper extended research done for a 2018 paper (Davoodian et al.) which focused primarily on Australian species.
The first problem with the 2020 paper is that it relies on too little phylogenetic data. Only one gene, rpb2 (RNA polymerase II, the second largest subunit), is sequenced, and justification for this minimalist methodology is not provided. The standard in 2020 was multilocus sequencing—something like three to seven genes, including at a minimum ITS, LSU, rpb2, and tef1 for mycorrhizal mushrooms. These days (writing in 2026) that standard persists but is clearly headed in the direction of full-genome sequencing. The reason multilocus sequencing is required by responsible mycology is that phylograms representing species can change from gene to gene, and using several genes helps to stabilize these changes and provide better support for the hypothesized species.
A second issue is the decision to call virtually every terminal clade in the phylogram a "species." Here is the relevant portion of the phylogram—the part representing what could be called "the castaneus clade":
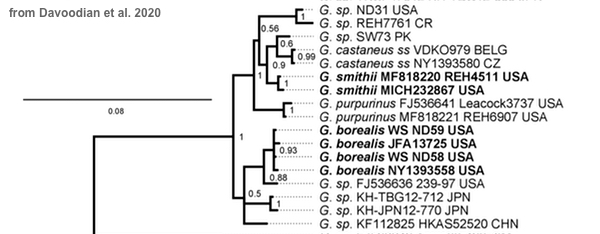
Note the scale bar, which represents 8% phylogenetic difference. If you divide it up (mentally) and superimpose it over the branches of the phylogram, you can see that all of the individual species branches in the group represent less than a two percent difference from their sisters, and some (for example castaneus and smithii) are not even one percent divergent. (By the way, the small numbers within the branching represent statistical support probabilities calculated by the software, not phylogenetic difference.)
There is no hard-and-fast, written-somewhere "rule" about this, but phylogenetic differences of less than two percent in one gene do not necessarily qualify as species lines for me. The standard I'm currently using for my own work on mycorrhizal mushrooms, using a minimum of four genes, is this:
"To delimit a species we required a minimum of three sampled collections in a monophyletic group with a minimum of 2% phylogenetic divergence measured as branch lengths, together with at least one of the following: a) inferred mycorrhizal host differences, b) unique geographic provenance, c) unambiguous, consistent morphological separation. We have only named new species when the material studied includes at least one of our own collections (including collections that were documented and sent to us by others)" (Kuo et al. 2026, ms. in prep.)
By this standard none of the putative species in the castaneus clade would be phylogenetically distant enough from castaneus itself to qualify as a separate species, and only borealis, with four collections tested, would have included enough collections even to merit consideration. Thus, while there are some geographic and morphological differences represented in the Davoodian et al. phylogram (purpurinus is purplish red, for example), my standards close the phylogenetic-difference door for all the branches in the phylogram, and leave the number-studied door open only for borealis, which still can't get through the first door.
Even if you feel my standard is too conservative and change the phylogenetic difference gateway to one percent (for context, humans and chimpanzees share 99% of DNA when compared gene-to-gene, but share about 96% when genomes are compared), castaneus and smithii would not qualify as separate, and borealis would have to include the unnamed collection it is sister to (a poorly documented GenBank sequence with no collection location, date, collector, or herbarium cited in the GenBank deposit or the original paper, Wilson et al. 2012). It is unclear whether the authors view the poorly documented GenBank sequence as a potentially discrete species; they say it "nests close to G. borealis" but, much to their credit, they continue: "however, since the physical specimen is unknown to the authors it is not treated here" (Davoodian et al. 2020). I wonder, then, why the sequence has been included in the phylogram in the first place.
Before I leave the phylogenetic issues I should also point out that Gyroporus castaneus is represented in the phylogram above by two GenBank sequences—a Belgian and a Czech collection, when the original description of the species represents French material (Bulliard 1788). There is no physical holotype collection made by Bulliard in the 18th century and available in a herbarium somewhere for sequencing, which means the species should be "epitypified" with a well documented contemporary collection from France for comparison—before potential species that are very closely related are considered.
Color
Gyroporus smithii and Gyroporus borealis can be separated, according to Davoodian et al. (2020), not just by sequencing the rpb2 gene and accepting very small phylogenetic divergence as division of species, but also on the basis of their colors, distributional range, and spore sizes:
"G. smithii corresponds to an orange species of Gyroporus that has often been encountered in the United States west and south of the Appalachian Mountains (east of the Great Plains) and is likely often mistaken for G. borealis (since G. borealis can display some orange coloration, though it is generally more darkly colored). G. smithii also has smaller and less elongated spores than G. borealis."
The problem is, these claims are supported by examination of a mere seven collections (four for borealis, three for smithii), and they fall apart with the slightest scrutiny. The color, for example, is described as "orange-brown, cinnamon brown to sometimes reddish" for borealis, and we are given the Methuen Handbook of Colour (Kornerup & Wanscher 1978) codes "6D–E6,5." The holotype collection is illustrated with a photo:
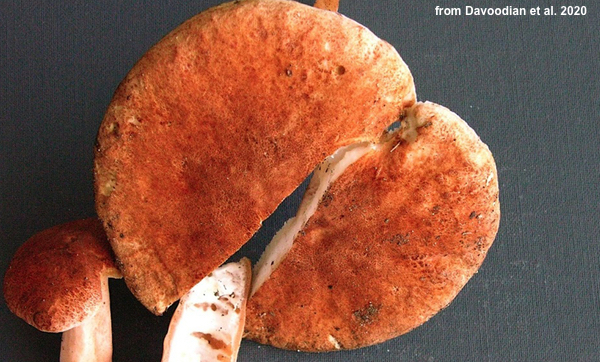
. . . that looks way more orange than "orange-brown." In fact, the Methuen codes given for borealis look like this:

These Methuen colors do appear to me to correspond to "orange-brown," but they don't match the type specimen, the Kuo color code for which would be RFO (O = "orange"). Four collections of borealis are cited, including this type collection. Two of the other three were made by Walt Sturgeon, one of the most experienced collectors on the planet. Sturgeon (2022, pers. com.) was asked for "specimens of the generally smaller orange capped one," and sent the two Ohio collections cited as paratypes. This means that three of the four collections were orange—but, remember, this is not the "orange species!"
By way of supposed contrast, the orange species, smithii, is described as "yellow-orange to orange to cinnamon-orange 6C–D7,6" (Davoodian et al. 2020). Here are these Methuen colors:

Yes, these colors are a bit more "orange" than those coded for borealis, although 6:D6 is an overlapping color. But even these versions of orange and brown are not as orange as the type collection of the non-orange species pictured above. I would offer you the potentially contrasting photo of one of the three smithii collections in the publication, BUT THERE IS NO PHOTO in the publication.
Add to all of this the potential for a single specimen to demonstrate the full range of Methuen colors illustrated above for both species in the course of its development, due to sunlight exposure, moisture levels, and so on . . . and we have arrived at what is essentially an exercise in faux science, along with an impossibly misleading and apparently erroneous separation of borealis and smithii on the basis of their cap colors.
Range
The epithet borealis, of course, means "boreal." The holotype collection of Gyroporus borealis (the very orange collection in the photo above) was made in Vermont—but the three paratype collections were made in northeastern Ohio and in Seattle, on the U-Dub campus. The Seattle collection is described as potentially aberrant, resulting from human transplant of ornamental eastern North American trees. So only one of the type collections is actually "boreal," and one wonders whether the authors meant "northern," which would still be a stretch for Ohio, but which ought to have been represented by the epithet septentrionalis . . . or whether the first author, based in Florida, just thought of anything north of Florida as "boreal," kind of the way my Chicagoland students used to think of anything south of Cook County and the collar counties as "southern Illinois."
Meanwhile the three collections cited for smithii are from Florida and southeastern Michigan (which is farther north than the Ohio collections); this apparently justifies stating the mushroom has "often been encountered in the United States west and south of the Appalachian Mountains (east of the Great Plains)." Three collections do not constitute "often," and confidently stating a range for either smithii or borealis on the basis of seven collections, one of which is aberrant, would result in a hasty, sustained objection on Law and Order for "assuming facts not in evidence."
Spore Size
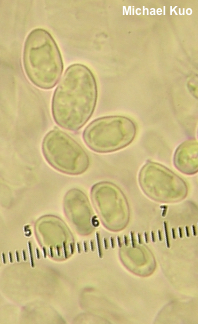
One needs to have compared a lot of spores from a lot of collections before declaring differences. The Davoodian paper (2020) decides that smithii and borealis can be separated on spore dimensions after examining seven collections. Spores of smithii measure "6–10 x 4–5 µm," while borealis spores are "(8.0–)8.8–11(–12) x 4.8–5.6(–6.4) µm." The parentheticals in the borealis numbers are meant to represent "outliers," according to the Materials & Methods section of the paper. With these measurements the authors conclude that "G. smithii . . . has smaller and less elongated spores than G. borealis." But take a look at this:
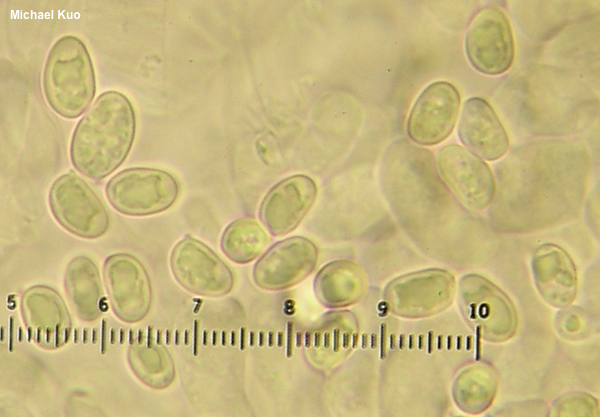
Spores here, in one of my collections from the castaneus group, range from 5.5 x 4 µm to 11.5 x 6 µm, and this variability is present in many of the 14 North American collections in my herbarium. Which species would you assign to this collection, based on the measurements? The length and width span the entire stated range for both species, including the supposed outliers.
This mistake, stating microscopic differences with confidence on the basis of insufficient data, is one I have made several times over the years—perhaps most egregiously when I named a morel Morchella capitata due to the microscopic "capitate elements on the sterile ridges" (Kuo et al. 2012) after examining a single holotype specimen. So I hope it is clear I do not intend to be righteous in these criticisms; I just want to establish with evidence and documentation that Gyroporus borealis and smithii, if you believe they qualify as distinct phylogenetic species, cannot actually be reliably separated with spore measurements—or with colors or distribution.
North American Gyroporus cyanescens Sloppiness
But wait, there's more. Sloppy papers abound in the Gyroporus cyanescens species group for North America, as well. This straw-colored, deeply bluing mushroom was assumed to be the same as cyanescens in Europe (first described from France by Bulliard, 1788) for more than a century—until a British mycologist visiting Michigan, Roy Watling (1969), described Gyroporus cyanescens var. violaceotinctus from the eastern arm of Michigan's Upper Peninsula, "differing markedly in the colour change of the flesh when it was exposed to the air; the flesh changed first lilac then rich purple and finally indigo." In contrast, the typical variety, corresponding to the European species, stained "greenish yellow when cut and finally azure blue, greenish blue in stipe rind and usually more cobalt blue in the pith." Watling believed that both varieties appeared in the Great Lakes region. Watling cited one additional Michigan collection for var. violaceotinctus, misspelling the collection location ("Pickney" for Pinckney), and three vaguely referenced North American collections for var. cyanescens.
Smith and Thiers (1971), however, found only var. violaceotinctus to be present in Michigan in their epic treatment of The Boletes of Michigan, describing its color change as "instantly indigo-blue when cut," with no mention of a lilac to purple stage; they cite 51 Michigan collections from across the state.
Fast forward to 2006. In a brief aside in a paper on "the sclerodermatoid fungi," Watling claims that var. violaceotinctus "has been shown by molecular studies to be more than 'skin-deep' and represents real speciation (Vigalys [SIC], personal communication)." Again, as above, this "assumes facts not in evidence." Or better said, the evidence is hearsay, since it is not presented, but merely attributed to the word of a well-known mycologist, Rytas Vilgalys, whose data is not presented and whose name is not even spelled correctly.
The first actually published North American cyanescens sequence that I find comes in Wilson et al. (2012), in a paper focused on "diversity and evolution of ectomycorrhizal host associations in the Sclerodermatinae." Here a poorly documented collection, "MB 05-001" (mislabeled "MB 05-004" in one of the phylograms) from "USA," shows clear phylogenetic separation from Australian cyanescens-group collections and from a poorly documented German collection (no collection date, no location beyond "Germany," no herbarium cited), possibly representing the "true" cyanescens in Europe. The North American collection was probably made by Manfred Binder ("MB"), one of the paper's coauthors, but the collection date, precise location within "USA," and the herbarium holding the collection are not provided; nor is this information available in the paper's supplementary materials, nor in the corresponding GenBank deposit. So we have no way of re-creating the sequencing experiment—and if the experiment cannot be repeated, it isn't science, as we all learned in 6th grade science class.
In 2015 Vizzini et al. use the Wilson et al. paper to support the idea that "in G. cyanescens cryptic speciation events have taken place," and put the GenBank sequence from Binder's "USA" collection in an LSU phylogram, where it is separated as sister, with low statistical support, to an Italian cyanescens collection. The Vizzini paper also provides an ITS phylogram in which three North American and five European cyanescens collections, including an Italian collection designated as an epitype, are minimally separated. Nevertheless, the paper keeps the sloppiness ball rolling by claiming that "[i]n particular, G. cyanescens var. violaceotinctus Watl. from North America is a real independent species (Vilgalys, personal communication, unpubl. data in Watling 2006)." I suppose spelling Rytas Vilgalys's name correctly represents a step forward, but citing a 19-year-old personal communication made to someone else, without providing any data, accomplishes at least two steps in the backward direction.
Just to wrap up the sloppiness package with a big sloppy ribbon, Blanco-Dios (2018) published the combination Gyroporus violaceotinctus for Watling's Michigan collection, saying only that "the known morphological and molecular data suggest that Gyroporus cyanescens is a complex of cryptic species (Vizzini et al., 2015); therefore the taxon designated as G. cyanescens var. violaceotinctus Watling, described from North America (Watling 1969) would be an independent species, as we propose here" (Blanco-Dios 2018, my translation) (oh, and there are no coauthors, so the "we" must be the royal we).
In the end all we can say with confidence is that someone needs to study a whole bunch of well documented North American Gyroporus cyanescens collections and stack them up in a multilocus phylogeny against European and global cyanescens group collections before we will know much. In other words, some old-fashioned, non-sloppy, thorough science needs to be done. However, since Watling's variety violaceotinctus was legitimately published, along with Blanco-Dios's elevation of the taxon to species level, Gyroporus violaceotinctus would be an appropriate name for any North American material that matched the type collection's DNA, if it were phylogenetically distinct from the European Gyroporus cyanescens.
References
Bessette, A. E., W. C. Roody & A. R. Bessette (2016). Boletes of eastern North America. Syracuse, NY: Syracuse University Press. 469 pp.
Blanco-Dios, J. B. (2018). Notas nomenclaturales en los órdenes Agaricales y Boletales. Tarrelos 20: 28–31.
Both, E. E. (1993). The boletes of North America: A compendium. Buffalo NY: Buffalo Museum of Science. 436 pp.
Coker, W. C. and Beers, A. H. (1943). The boleti of North Carolina. New York: Dover. 96 pp. (1971 reprint.)
Davoodian, N. & R. E. Halling (2013). Validation and typification of Gyroporus purpurinus. Mycotaxon 125: 103–105.
Davoodian, N. & 14 coauthors (2018). A global view of Gyroporus: molecular phylogenetics, diversity patterns, and new species. Mycologia 110: 985–995.
Davoodian, N., K. Hosaka, O. Raspé, O. A. Asher, A. R. Franck, A. de Kesel, T. P. Delaney, J. F. Ammirati, E. Nagasawa, B. Buyck & R. E. Halling (2020). Diversity of Gyroporus (Gyroporaceae, Boletales); rpb2 phylogeny and three new species. Phytotaxa 434: 208–218.
Grund, D. W. & Harrison, A. K. (1976). Nova Scotian boletes. Germany: J. Cramer. 283 pp.
Kornerup, A. & J. H. Wanscher (1981). Methuen handbook of colour. 3rd edition. London: Methuen & Co., Ltd. 252 pp.
Murrill, W. A. (1914). American boletes. New York: W. A. Murrill. 40 pp.
Ortiz-Santana, B., D. J. Lodge, T. J. Baroni & E. E. Both (2007). Boletes from Belize and the Dominican Republic. Fungal Diversity 27: 247–416.
Noordeloos, M. E. (2018). Gyroporaceae. In Noordeloos, M. E., T. W. Kuyper, I. Somhorst & E. C. Vellinga, eds. Flora Agaricina Neerlandica: Critical revisions of families of agarics and boleti occurring in the Netherlands. Volume 7. Orrigio, Italy: Candusso Editrice. 185–188.
Siegel, N. & C. Schwarz (2016). Mushrooms of the redwood coast: a comprehensive guide to the fungi of coastal northern California. Berkeley: Ten Speed Press. 602 pp.
Singer, R. (1945). The boletinae of Florida. Germany: J. Cramer. 126 pp. (1977 reprint.)
Smith, A. H. & Thiers, H. D. (1971). The boletes of Michigan. Ann Arbor: U Michigan P. 428 pp.
Snell, W. H. & Dick, E. A. (1970). The boleti of northeastern North America. Germany: J. Cramer. 115 pp.
Tedersoo, L., T. W. May & M. E. Smith (2010). Ectomycorrhizal lifestyle in fungi: global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 20: 217–263.
Vizzini, A., C. Angelini & E. Ercole (2015). Molecular confirmation of Gyroporus lacteus and typification of Boletus cyanescens. Phytotaxa 226: 27–38.
Watling, R. (1969). New fungi from Michigan. Notes from the Royal Botanic Garden, Edinburgh 29: 59–66.
Watling, R. (2006). The sclerodermatoid fungi. Mycoscience 47: 18–24.
Wilson, A. W., E. A. Hobbie & D. S. Hibbett (2007). The ectomycorrhizal status of Calostoma cinnabarinum determined using isotopic, molecular, and morphological methods. Canadian Journal of Botany 85: 385–393.
This site contains no information about the edibility or toxicity of mushrooms.
Kuo, Michael (2026, June). Gyroporus. Retrieved from the Mushroomexpert.Com website: www.mushroomexpert.com/gyroporus.html
All text and images © , Mushroomexpert.com