|
|

|
Glossary |
|
Adnate Gills, Adnexed Gills See gills. |
|
|
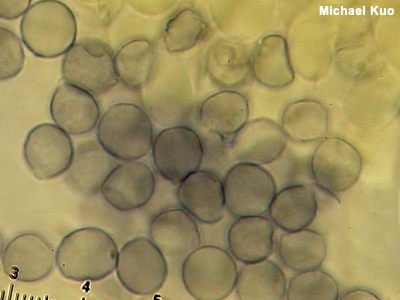
Amyloid An amyloid reaction is a bluish-black color change when something is mounted for the microscope in an iodine-based reagent like Melzer's Reagent or Lugol's Solution. Spores are typically what is looked at to determine whether the reaction is amyloid or not—but other microscopic structures sometimes demonstrate the color change, too. Figuring out what genus your mushroom belongs in is sometimes made easier by knowing whether or not it has amyloid, inamyloid, or dextrinoid spores. Examples include Rhodocollybia versus Gymnopus (dextrinoid versus inamyloid spores) and Porpoloma versus Tricholoma (amyloid versus inamyloid spores). Identification in the genus Amanita is often facilitated by figuring out whether a collection has amyloid or inamyloid spores. Mycenoid mushroom identification sometimes relies on whether spores are amyloid or not, as well. The frequently beautiful ornamentation of spores in Lactarius and Russula is amyloid, often strongly so, which is why the ornamentation is so visible (and why microscopy in these genera requires a Melzer's mount). Compare with dextrinoid and inamyloid. See also Using a microscope: viewing and measuring spores. |
|
|
Anamorph, Anamorphic The life cycle of some fungi involves both sexual and asexual stages. For such fungi the anamorph is the asexual stage, while the teleomorph is the sexual stage. In the anamorphic stage asexual reproduction may occur through cloning, with the production of conidia (see spores for more information). In older taxonomic schemes anamorphs and teleomorphs were sometimes placed in different genera, despite being life cycle stages of the same organism. For example, the anamorph to the left was often recognized as "Xylocoremium flabelliforme," while its teleomorph was Xylaria cubensis. However, more current taxonomic rules disallow this practice, and the name Xylocoremium flabelliforme is now a deprecated synonym for Xylaria cubensis. Anamorphs are sometimes called "imperfect fungi." Compare with teleomorph. |

|
|
Annulus, Ring A ring of tissue around the upper part of a mushroom's stem, resulting from the collapsing of the partial veil, is an "annulus" in Mycologese—or just a "ring" in plain English. Rings are extremely variable, ranging from ephemeral and quickly disintegrating to sturdy and prominent. The type of ring a mushroom has is frequently a key feature for identification. Some mushrooms, like Macrolepiota procera, have rings which are separable from the stem and can be made to slide up and down. Other rings may be peronate (sheathlike, like the ring of Agaricus bitorquis), pendant (skirtlike, like the ring of Amanita magnivelaris), or flaring (like the ring of Tricholoma caligatum, illustrated to the left). Additionally, the position of the ring can be important; it may be superior, apical, median, inferior, or basal ("at the very top" to "at the very bottom," in order). Rings are notoriously absent when they should be present, so be sure to have both mature and immature specimens on hand when attempting difficult identifications; the partial veil will show up more clearly with immature specimens. Compare with ring zone. |

|
|
Appendiculate An appendiculate cap is one in which the tissue of the universal veil hangs over the edge after the cap has expanded and ruptured the veil, as in the illustration of Amanita daucipes to the left. In the genus Amanita, especially, identification keys often ask for an assessment of whether the cap is appendiculate or not, but the term is occasionally found elsewhere in the mushroom world. The feature is not as stable as one might like, however, due to environmental factors like rain and wind, as well as the fact that mushrooms do not read identification keys, so it is not uncommon to find a mushroom that "should" have an appendiculate cap but does not. See also universal veil. |

|
|
Areolate "Areolate," in a mushroom mycology context, means "cracked," usually in age, like many of us. The cap surface of just about any mushroom can become cracked in dry weather conditions, but some species typically develop cracked caps in normal weather conditions. Xerocomellus chrysenteron is a well-known example. Sometimes the color of the flesh, revealed between the cracks of an areolate cap, is also an important identification feature. Because the "crackedness" of a mushroom's cap is so dependent on environmental conditions, I do not recommend stressing it too heavily in your identification decisions—especially among the boletes, which are notorious for cracking up when they shouldn't. What they think is so damned funny is beyond me. |

|
|
Ascus, Asci, Ascomycota An ascus (plural asci, which is honest-to-God pronounced "ass eye" in American English) is a microscopic structure in which spores are produced. Asci cover the spore-bearing surfaces of many mushrooms; these mushrooms are consequently members of the phylum Ascomycota. Spores are forcibly ejected from asci at maturity—often, simultaneously across the spore producing surface, creating a puff of spore "smoke" that can be visible to the naked eye—and even, in some cases, producing a hissing sound! The number of spores held in a mushroom's asci (typically 8, for species of interest to most mushroom hunters, but sometimes 4, 6, and so on) can occasionally be an informative character in identification choices. A more commonly needed microscopic assessment, however, involves whether the tips of the asci are amyloid and thus turn blue in Melzer's reagent, like the asci of Peziza species like Peziza michelii, pictured to the left. Compare with basidium, basidia, basidiole, Basidiomycota. See also Phylum: Ascomycota on the taxonomy page, paraphyses, and using a microscope. |
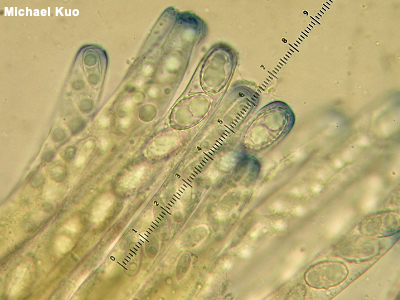
|
Basal Mycelium See mycelium. |
|
|
Basidium, Basidia, Basidiole, Basidiomycota A basidium (plural basidia) is a microscopic structure on which spores are produced. Basidia cover the gills, tubes, or other spore-bearing surfaces of many mushrooms; these mushrooms are consequently members of the phylum Basidiomycota. Basidia typically develop apical prongs ("sterigmata") on which spores develop. Determining the number of prongs on a mushroom's basidia (usually 2 or 4) can sometimes be useful for identification (for example in Craterellus or Amanita). At maturity spores are flung from the basidium's prong into the air currents. Basidioles are sterile, basidium-like structures—meaning they do not produce spores and do not have prongs. Compare with ascus, asci, Ascomycota. See also Phylum: Basidiomycota on the taxonomy page, and using a microscope. |
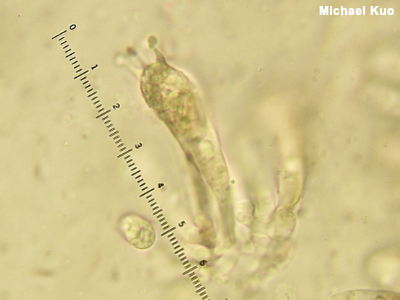
|
Basionym See protologue. |
|
Binding Hyphae See hyphae. |
|
|
Biological Species, Biological Species Concept A biological species is a species defined using the concept commonly accepted and applied (in the popular mind, anyway) in zoology: if animals can mate and produce offspring, they are the same species. With mushrooms, however, "mating" is much more complicated. For our purposes here, suffice it to say that mycologists use petri-dish cultures of mushrooms and attempt to pair them with other cultures in mating studies in order to determine whether they constitute the same species. Disturbingly, mycologists have discovered the possibility that some mushrooms may retain the ability to mate—perhaps as some sort of ancestral vestige—despite having diverged as phylogenetic species. Pleurotus pulmonarius, illustrated to the left, is well-supported as a biological species. It also makes a good morphological species and a good phylogenetic species, so all of its species-concept-ducks are in line. Compare with morphological species and phylogenetic species. |

|
Boletinoid Pores See pores, pore surface. |
|
|
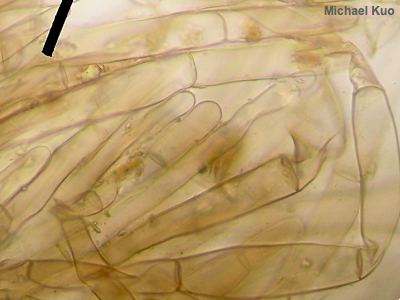
Brachybasidioles, Pavement Cells Brachybasidioles are microscopic structures present in some gilled mushrooms—particularly those that are short-lived, like species of Bolbitius and some coprinoid mushrooms, which can appear and collapse within a few hours of morning sunlight. Brachybasidioles appear between basidia, almost as though their function is to prop up the spore-bearing structures. In the illustration to the left, the round-looking items are basidia, while the squarish, blocky structures separating the basidia are the brachybasidioles (the reddish brown, out of focus things are spores). You are looking down on the spore producing surface from above, so you're seeing the tops of the structures—and getting a good view of why brachybasidioles are also called "pavement cells." See also using a microscope. |
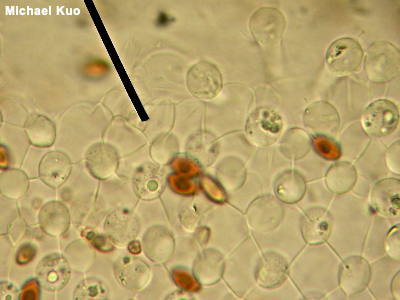
|
|
Brown Rot The mycelia of brown rot fungi degrade cellulose in the wood they inhabit, resulting in a characteristic decay of the wood, which turns brown and breaks up into more or less cubical chunks. Brown rot fungi are especially common on the wood of conifers, but can be found on hardwoods as well. Well-known brown rot fungi include species of Laetiporus, Gloeophyllum sepiarium, and Phaeolus schweinitzii. Compare with white rot. |

|
|
Butt Rot The mycelia of butt rot fungi attack trees through their roots and produce decay in the root system and the heartwood of the lower portion of the tree (up to about 10 or 20 feet above ground). The result is a weakened, or even hollow, tree base—which makes the tree more susceptible to windthrow, especially if other agents (beetles, for example) have combined destructive forces with the fungus. The mushrooms produced by butt rot fungi are almost always positioned near the base of the tree, either fruiting from the main trunk or appearing terrestrial. Many polypores are butt rot fungi, including the well-known species Inonotus dryadeus, Heterobasidion anosum, Bondarzewia berkeleyi, and Phaeolus schweinitzii. |

|
Caulocystidia See cystidia. |
|
Cheilocystidia See cystidia. |
|
Chrysocystidia See cystidia. |
|
|
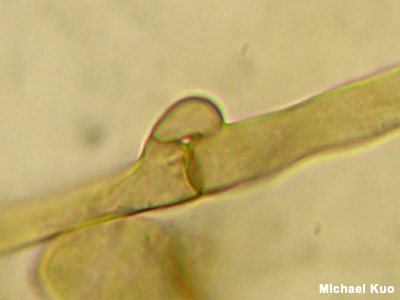
Clamp Connection A clamp connection is a connection between two hyphae (fungal cells). Rather than merely terminating with a septum (a simple dividing wall), clamped cells involve a little arm, or clamp, that reaches from one cell to the next, appearing to hold the cells together. Some mushrooms have clamp connections; others don't. Determining whether clamp connections are present in your mushroom will certainly help you make progress through many traditional, microscope-based identification keys. But it must be pointed out that recent DNA-based research has called into question the whole idea of whether clamp connections are always "informative," consistent characters. Cantharellus and Craterellus, for example, used to be sorted out in large part on the basis of clamp connections—but DNA has shown us that our arrangements of these genera based on clamps did not provide an accurate picture of what is related to what, and that clamp connectionss, in fact, don't seem to have any correlation to phylogenetic groupings. Assessing whether basidia have clamp connections at their bases can occasionally be important for identifying mushrooms—for example in Armillaria, where the basidia of Armillaria mellea are not basally clamped as they are in other species. See also using a microscope. |
|
Conidia See spores. |
|
|
Cortina A cortina is a form of partial veil consisting of a cobweb-like protective covering over the immature spore bearing surfaces. Cortinas are variable; they can be thin and arachnoid, consisting of a few spider-web-like threads—or they can be thicker and more dense (sometimes so thick and dense that the line between cortina and ring can be blurred). Sometimes cortinas can be very difficult to see; make sure you are examining very young specimens, and use a hand lens! Cortinas typically stretch open and fall apart as the mushroom matures, disappearing entirely or leaving a ring zone on the stem. As mushrooms mature and develop spores, still-attached cortinas sometimes catch spores and appear to have changed color as a result, as in the illustration to the left, wherein the far-left cortina has been covered with rusty brown spore dust. The genus Cortinarius is so named because its members typically have a cortina when young. Other genera that often feature cortinas include Hebeloma and Inocybe. A few tricholomas and waxy caps feature cortinas; for example the presence or absence of a cortina when in the button stage can help separate Hygrophorus erubescens from Hygrophorus purpurascens. Compare with annulus, ring. See also partial veil and ring zone. |

|
Cryptic Species See phylogenetic species concept. |
|
Cutis See pileipellis. |
|
|
Cystidium, Cystidia Cystidia (singular: cystidium) are special, sterile cells viewed under the microscope. The presence or absence of cystidia—as well as their shapes and sizes, if they are present—is sometimes important in mushroom identification. Cystidia can appear anywhere on a mushroom's fruiting body. On mushrooms with gills, cystidia on the edges of the gills are called cheilocystidia, while cystidia on the faces of the gills are called pleurocystidia. On mushrooms with tubes, cheilocystidia appear on the mouths of the tubes while pleurocystidia are on the tube walls. Cystidia on a mushroom's stem are caulocystidia; cystidia on a mushroom's cap are pileocystidia. Chrysocystidia are cystidia with yellowish refractive contents. Metuloids are prominently projecting pleurocystidia with thick walls. Skeletocystidia are the thick-walled, cystidium-like ends of skeletal hyphae that project through the spore-bearing surface. "Cystidioles" are small cystidia that do not project much beyond the basidia. See also setae and using a microscope. |
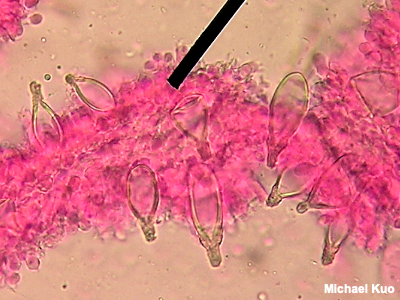
|
|
Dextrinoid A dextrinoid reaction is a reddish brown color change when something is mounted for the microscope in an iodine-based reagent like Melzer's Reagent or Lugol's Solution. Spores are typically what is looked at to determine whether the reaction is dextrinoid or not—but other microscopic structures sometimes demonstrate the color change, too. Figuring out what genus your mushroom belongs in is sometimes made easier by knowing whether or not it has amyloid, inamyloid, or dextrinoid spores. Examples include Rhodocollybia versus Gymnopus (dextrinoid versus inamyloid spores) and Porpoloma versus Tricholoma (amyloid versus inamyloid spores). The genus Rhodocollybia can be separated from other collybioid mushrooms on the basis of its dextrinoid spores. Lepiotoid mushrooms often have dextrinoid spores, as do some marasmioid mushrooms, along with species of Cortinarius, Galerina, Gymnopilus, and Hebeloma, among others. Compare with amyloid and inamyloid; see also Using a microscope: viewing and measuring spores. |
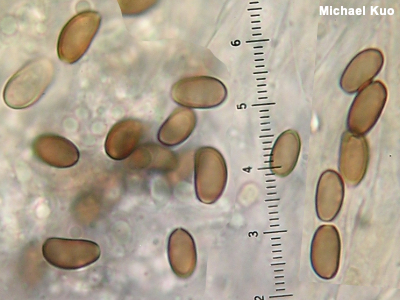
|
Decurrent Gills See gills. |
|
Dimitic Hyphal System See hyphae. |
|
Ectomycorrhizal, Ectomycorrhiza/ae See mycorrhizal. |
|
|
Effused-Reflexed An effused-reflexed mushroom is one that is closely appressed to the substrate (resupinate) except for a small margin that extends to form a rudimentary cap-like structure. Effused-reflexed mushrooms are found among crust fungi and polypores. For many mushrooms, the "decision" to develop an effused-reflexed fruiting body or a truly pileate fruiting body is simply a matter of where the mushroom is growing (on the underside of a log, on its side, or on top of the log) and what fruiting body style will best expand the spore-producing surface. Ischnoderma resinosum, for example, is equally happy being resupinate, effused-reflexed, or pileate. Compare with resupinate and pileate. |

|
Endomycorrhizal, Endomycorrhiza/ae See mycorrhizal. |
|
Epithelium See pileipellis. |
|
|
Excipular Surface / Sterile Surface In cup fungi and in other ascomycetes the term "excipular surface" refers to the sterile surface opposite the hymenium, or fertile surface, where the spore-producing machinery is located. Compare with hymenium. |
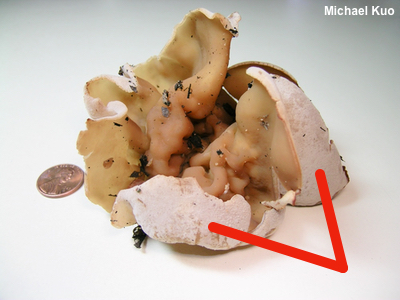
|
Fairy Ring See the page for fairy rings. |
|
|
False Gills False gills are folds in a mushroom's spore-bearing surface that can approximate the appearance of gills, but differ by not being structurally distinct units. False gills appear in the chanterelles and in a few other mushrooms. Assessing whether gills are "false" or not is not always easy, since some Cantharellus species can develop very gill-like false gills. But if you have carefully examined the gills on truly-gilled mushrooms (for example the common store-bought mushroom, Agaricus bisporus), you will have noticed that each gill is structurally separate; you can, for example, separate it from the cap fairly easily, and it doesn't seem as though it was part of the cap. Separating a false gill from the mushroom, however, is not as easily accomplished, since the interior of the false gill is actually composed of the mushroom's flesh. Compare with gills, lamellae. |

|
Friable See volva. |
|
Generative Hyphae See hyphae. |
|
|
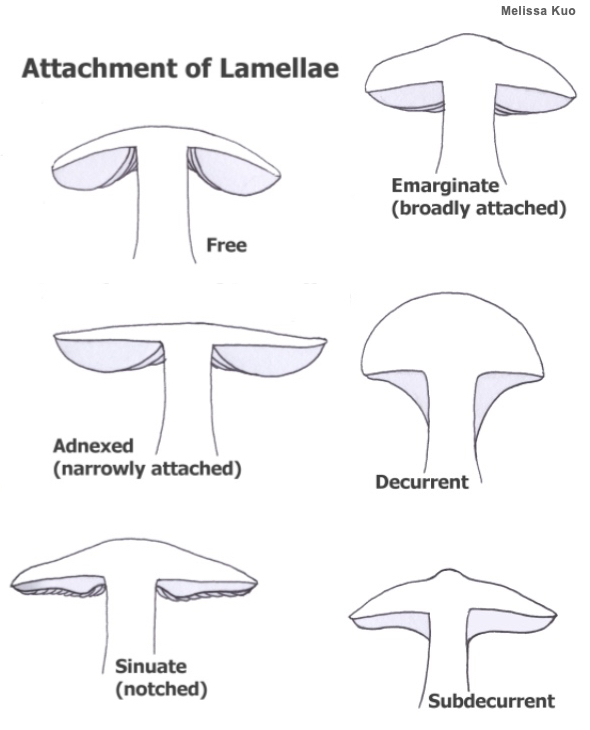
Gills, Lamella, Lamellae Gills ("lamellae" in Mycologese) are plate-like or blade-like structures attached to the underside of the cap in many members of the Basidiomycota. Mushrooms with gills are commonly called "gilled mushrooms," but that term has little scientific meaning since gills developed several times on the evolutionary tree and the presence of gills does not, therefore, necessarily signify close relationship. Gills are covered with spore-producing basidia, and represent an ingenious way to expand the mushroom's spore producing surface; imagine the total surface area of all the gills on a mushroom (both sides!), as compared to the surface area of a single, flat surface the size of the mushroom's cap. Assessing the morphology of a mushroom's gills is crucial in mushroom identification. Gill attachment to the stem and gill spacing are illustrated below. Other important gill details include their color (which can change as the mushroom matures) and consistency (brittle, for example, or waxy), as well as the presence or absence of short gills. Some mushrooms have gills with serrated edges (for example, species of Lentinellus). Other mushrooms have frequently forked gills (Russula variata), or gills with cross-veins (Xeromphalina kauffmanii). Compare with false gills and short-gills. See also "What, if anything, is a gilled mushroom?" on the page for Lenzites betulinus.
|


|
|
Glandular Dots Glandular dots are aggregations of small, pigmented cells that appear to the naked eye like dots. Glandular dots are found in Suillus. The dots are usually very small, and result from clusters of pigmented, inflated cells on the stem surface. Identifying Suillus species often hinges on the presence or absence of glandular dots—but this can be a frustrating character to assess, since many species have whitish or pale glandular dots that do not darken and become conspicuous until maturity, or when the mushroom is dried. Compare with scabers. |

|
|
Gleba, Glebifer The term "gleba" is used in two fairly distinct ways: 1) In puffballs, the spore-producing portion of the interior is called the gleba. Typically a puffball's gleba is fleshy at first and then disintegrates, becoming dusty spore powder. In some puffballs the gleba is distinct from a sterile base, which does not turn into spore dust. Puffball gleba is illustrated in the glossary entry for sterile base. 2) In stinkhorns, gleba is the stinky, sticky, usually-olive-or-brown, spore-filled slime that covers some surface or surfaces on the stinkhorn to attract insects for spore dispersal. In the illustration to the left, dark brown gleba covers the head of Phallus impudicus. A glebifer is the organ that produces gleba on a stinkhorn. Concerning puffballs, compare with sterile base. |

|
Host-Specific See mycorrhizal. |
|
|
Hygrophanous A mushroom cap that changes color markedly as it dries out—often resulting in a two-toned appearance during the process—is "hygrophanous." Well-known mushrooms that often demonstrate hygrophanous caps include Galerina marginata (illustrated to the left), Panaeolus foenisecii, and Psathyrella candolleana. In the genus Cortinarius, assessing whether the cap is hygrophanous can be important in narrowing down identification choices, since the traditional subgenus Telamonia is (partly) separated on the basis of the hygrophanous caps of the species. There once was a chicken named Darren That's something you won't get anywhere else, ladies and germs. Mushroom. Expert. Dot. Com. |

|
Hymeniform See pileipellis. |
|
|
Hymenium A mushroom's hymenium is the surface on which it produces spores. Microscopic spore-producing structures (either asci or basidia) cover the hymenium and, at maturity, release spores. The hymenium of some mushrooms is flat and simple—as in cup fungi like Sarcoscypha austriaca, pictured to the left, or in the crust fungi. Other mushrooms, presumably in a strategy to increase surface area for the spore-producing machinery, create wrinkles, pockets, and folds (for example, morels). Even more spore-producing surface area is accomplished with tubes or gills. See also spores, tubes, gills, and using a microscope. Compare with excipular surface. |

|
|
Hypha, Hyphae, Hyphal System, Hyphal Peg, Hyphal Tower Hyphae (singular: hypha) are fungal cells; they are tube-like and elongated. For some mushrooms (primarily polypores), mycologists describe the types of hyphae that make up a mushroom, constituting the the "hyphal system." Generative hyphae are thin-walled, septate, branched hyphae, and are the fundamental type from which the other hyphal types arise. Skeletal hyphae are thick-walled, aseptate, and unbranched. Binding hyphae are thick-walled, aseptate, and distinctly branched. Monomitic hyphal systems are composed of generative hyphae only. Dimitic hyphal systems are composed of generative and skeletal hyphae, or are composed of generative and binding hyphae. Trimitic hyphal systems are composed of all three hyphal types. Hyphal pegs are found in some mushrooms (for example Lentinus tigrinus), and consist of small, peg-like aggregations of hyphae protruding from the hymenium. The Hyphal Tower, the world's tallest mushroom, is in Paris. See also using a microscope. |
|
Imperfect Fungi See anamorph. |
|
|
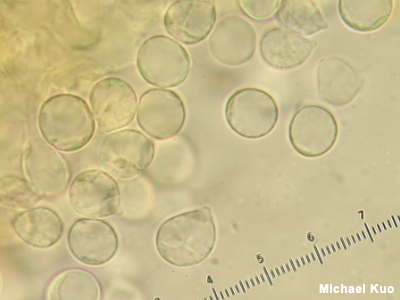
Inamyloid An inamyloid reaction is a negative (lack of) color change when something is mounted for the microscope in an iodine-based reagent like Melzer's Reagent or Lugol's solution. Spores are typically what is looked at to determine whether the reaction is inamyloid or not. Figuring out what genus your mushroom belongs in is sometimes made easier by knowing whether or not it has amyloid, inamyloid, or dextrinoid spores. Examples include Rhodocollybia versus Gymnopus (dextrinoid versus inamyloid spores) and Porpoloma versus Tricholoma (amyloid versus inamyloid spores). Identification in the genus Amanita is often facilitated by figuring out whether a collection has amyloid or inamyloid spores. Mycenoid mushroom identification sometimes relies on whether spores are amyloid or inamyloid, as well. Compare with dextrinoid and amyloid; see also See also spores, tubes, gills, and using a microscope: viewing and measuring spores. |
|
Ixocutis See pileipellis. |
|
Ixotrichoderm See pileipellis. |
|
|
KOH, Potassium Hydroxide KOH is the chemical symbol for potassium hydroxide—a strong base often used to study mushrooms. Although it is sometimes difficult to obtain, KOH can usually be purchased without too much difficulty. Several major online vendors have it available. KOH is used in a 2 percent aqueous solution as a mounting medium for microscopic examination of mushrooms. As a medium it often does a good job of clarifying mounts and making tissues and structures visible. It has its drawbacks (for example it tends to swell some structures) but it has been used for so long by mycologists that using it is necessary if one wants to compare data with their work.
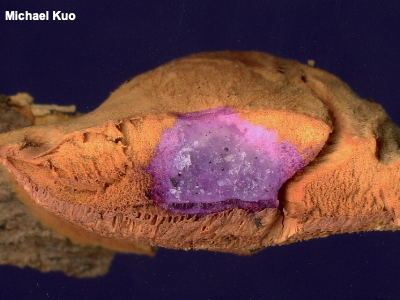
Away from the microscope, a stronger solution of KOH (somewhere in the neighborhood of 3–10 percent) is used to test chemical reactions on mushrooms' surfaces; color-change reactions like the dramatic purple on the flesh of Hapalopilus nidulans pictured to the left can be useful in identification. |
|
Lamellae, lamella See gills. |
|
Limb, Limbate See volva. |
|
Lugol's Solution Lugol's solution is an iodine-based stain that is often proposed as an easy-to-obtain substitute for Melzer's reagent (which is used to determine whether spores and tissues are amyloid, inamyloid, or dextrinoid). However, a study by Leonard (2006) in which "[t]he spores of 35 species of fungi were tested with Melzer's, Lugol's, and iodine solutions" found that "[a]ll 35 species reacted as predicted from authoritative sources with Melzer's but results were inconsistent with Lugol's and iodine." Compare with Melzer's reagent. |
|
|
Marginate In our context, marginate is usually a description for gills in which the edges are colored differently than the faces— as in Pluteus atromarginatus, to the left, which is named for its black-marginate (atro-marginatus) gills. Marginate gills are usually caused by microscopic cheilocystidia on the gill edges. Well-known mushrooms with marginate gills include Entoloma serrulatum and Mycena leaiana The term is also used occasionally to describe stem bases that have large bulbs with a flattened upper edge—for example Amanita abrupta or Agaricus reducibulbus. |

|
Mating Studies See biological species concept. |
|
|
Melzer's Reagent Melzer's reagent is an iodine-based stain regularly used in mycological microscope work to better see tissues and to determine whether spores and tissues are amyloid, inamyloid, or dextrinoid. It is unfortunately extremely difficult to obtain. Melzer's contains water, iodine, and potassium iodide, all of which are fairly easy to get hold of—but it also contains chloral hydrate, which is a controlled substance. Thus, you won't be able to buy it easily. Virtually your only option is to beg it from a professional mycologist. Even mycologists have difficulty obtaining Melzer's, however, and if the mycologist you know can't afford to provide you with some of her precious supply (or if you cannot find a mycologist), your last resort is to try explaining your situation to your doctor and getting a prescription for chloral hydrate (not likely; it's a date-rape drug) or for Melzer's reagent itself, which a compounding pharmacist could mix according to the formula below, and which your doctor would need to write on the prescription (still not very likely, but not unheard of). Water: 20.0 gm See also Lugol's solution. |

|
Metuloids See cystidia. |
|
Monomitic Hyphal System See hyphae. |
|
|
Morphological Species, Morphological Species Concept A morphological species is defined using the concept used through centuries in mycology, until fairly recently: if the mushrooms have significantly different morphology, they represent different species. An organism's morphology is its set of observable features, which can be macroscopic and/or microscopic. There are obvious problems with using morphology to define species. Where do you draw the lines? There are differences between individual mushrooms, of course; when do you decide that a group of mushrooms shares enough observable features to constitute a group we should label a "species?" How much importance do you attach to each of the various features? Is color more important than spore size? These questions (and many, many others) lead to an uncomfortable conclusion: it is scientists who determine what makes a species, rather than the mushrooms. In other words, there is nothing inherently natural about a morphological species; it represents merely what the human eye can see (with and without the help of technology). Lactarius gerardii, pictured to the left, was originally described in the 19th century as a morphological species. However, DNA research indicates there are several phylogenetic species within the morphological concept of "Lactarius gerardii." Compare with biological species and phylogenetic species. |

|
Mushroom Every so often I get an email asking me if the thing in the attached photo is a mushroom or a toadstool. I want to say: What the fuck does that even mean? But I don't. I just click "delete" and move on with my life. "Mushroom" isn't a scientific term. I once had a very good teacher who defined poetry as "any text someone asks you to consider as poetry." I'm inclined to do more or less the same with "mushrooms." They are fungi, but they are definitely not all fungi (a category that would include, for example, ring worm and the bluish mold that appears on your bread)—so where you want to draw the "mushroom" line within the fungi is up to you. That said, one good way of understanding mushrooms, I think, is to consider them as spore factories. Most of the time, the fungal organism in question spends its time as vegetative mycelium, looking nothing like the "mushroom" thing we're talking about. But when the reproductive urge takes hold (that's called "personification"), the fungus produces a spore factory—a structure to make babies and send them out into the world. Perhaps the organism designs a stem or a pseudostem, preparing to lift the spore-producing shop floor high enough so that the spores will easily catch air currents. Perhaps a cup is created, or a cap, to hold the floor. Asci or basidia, the actual spore-producing machines, are erected all across the floor. Some organisms increase the factory capacity immensely by increasing the floor size with gills, false gills, or tubes. Sometimes the factory adds safety features: veils are erected to cover and protect the machinery until the spores are ready. As soon as the spores are released, the factory shuts down. The "mushroom" wilts, decays, and eventually disappears. But the organism itself continues, perhaps in the same place, if the mycelium has not run out of nutrients—or somewhere else, when spores land in just the right places, germinate, and develop into mycelia. Pictured to the left is probably the most iconic "mushroom" in the world, Amanita muscaria var. muscaria. The Mario Kart mushroom. The old-farm-woman-bending-over-in-her-polka-dotted-dress mushroom. The hookah-smoking-caterpillar-Lewis-Carroll mushroom. Amanita muscaria is one of the more complex and elaborate (and beautiful!) spore factories; it features a long stem, a universal veil to protect the construction of the factory, a partial veil to protect the spore-producing machinery, and the perfect timing required to raise the cap up and expand it to break the veils and release spores. |

|
|
Mycelium A "mushroom" is only the reproductive part of the organism—in the way an apple is the fruit of the whole organism, the apple tree. The main part of a mushroom is underground, or running through the leaves or deadwood (or other substrate); it consists of a mass of hyphae, often visible to the naked eye as a whitish mass or as whitish threads; the mycelium of Marasmius delectans, spreading through hardwood leaf litter beneath the mushrooms, is illustrated to the left. Perhaps you have seen decaying leaves covered with whitish, fuzzy material before; in many cases this is the mycelium of a mushroom. The mycelium is present even when there are no mushrooms. In fact, mushrooms spend most of their life cycle as mycelium, vegetating and consuming nutrients from whatever substrate they inhabit, only producing what we know as "mushrooms" when it's time to reproduce. Some mushrooms, like species of Armillaria, have mycelia that can stretch for miles and miles; you may remember the recent discovery through DNA testing of the "world's largest organism," an Armillaria species in the Pacific Northwest. Basal mycelium: Evidence of the mycelium is sometimes found at the base of a mushroom's stem; there may be mycelial down or fuzz where the stem meets the ground. The presence or absence of basal mycelium is sometimes important in the identification process, as is its color. Frustratingly, current literature for both Laccaria and Phylloporus relies heavily on the color of the basal mycelium when the mushroom is fresh. In Laccaria one is asked to assess whether or not the basal mycelium was white or lilac—but the lilac colors are often faint to begin with, and fade very quickly. In Phylloporus the choice is yellow or white, and only slightly easier to determine.
Rhizoids, rhizomorphs: Mycelial cords or strands, called "rhizoids" or "rhizomorphs," are found in some species, and their presence sometimes helps in identification decisions. See the page for Armillaria mellea for an illustration of its large, black, conspicuous rhizomorphs, and see the illustrations on the page for Stropharia hardii for smaller, more typical, white rhizomorphs. Mycelium is not always as obvious as it is in the Marasmius photo to the left; to view the basal mycelium of a mushroom you must be sure to include the stem base when you pick it—and digging around in the substrate with a pocket knife is often necessary to find rhizomorphs. |

|
|
Mycorrhizal, Mycorrhiza Mushrooms that are mycorrhizal are involved in a symbiotic (mutually beneficial) relationship with the tiny rootlets of plants—usually trees. The hyphae of the mushroom's mycelium surround the tree rootlets with a sheath (called a mycorrhiza), and the mushroom helps the tree absorb water and nutrients while the tree provides sugars and amino acids to the mushroom. The organisms may need each other to survive. (Strictly speaking, the mushrooms I'm calling "mycorrhizal" are ectomycorrhizal. There are also endomycorrhizal fungi; their hyphae actually penetrate the plant's rootlets, rather than surrounding the rootlets with a sheath. However, endomycorrhizal fungi do not produce mushrooms.) Not all trees are mycorrhizal. Maples, for example, do not form (ecto)mycorrhizal relationships with fungi, while oaks and pines (among many others) are eager to partner with mushroom-producing fungi. Some mycorrhizal mushrooms are very host specific and will only form mycorrhizae with a certain species of tree. Suillus lakei is an example; it partners only with Douglas-fir. Other mycorrhizal mushrooms are willing to grow with closely related trees—for example, Suillus tomentosus will grow with two-needled pine species like lodgepole pine or jack pine. Others are less picky; Tylopilus rubrobrunneus will associate with many different oaks and hickories—and some mycorrhizal mushrooms are generalists, not very picky at all, willing to associate with hardwoods and conifers alike (for example, Amanita jacksonii). Mycorrhizal mushrooms can appear at quite a distance from the trees they are associated with, since the tree's tiny rootlets can extend well past the tree's drip-line. "Toppling range" may be a better determiner than the drip-line: if the tree were to topple over, some part of it, including the very top, could fall on the mushrooms. This range is farther than collectors usually consider for mycorrhizal association, and can easily be 80 feet or more for mature trees (or even hundreds of feet, if West-Coast Douglas-fir is considered). But I have seen clear evidence many times that "toppling" distance is not outside of mycorrhizal range—for example with Suillus americanus, appearing in my yard 80 feet away from its host, the only eastern white pine within half a mile; the tree was about 90 feet tall. Many of our most well-known mushrooms are mycorrhizal, including species of Amanita, Russula, Lactarius, Cortinarius, and most boletes. |

|
|
Paraphyses Paraphyses (singular: paraphysis) are sterile structures in the hymenium of some mushrooms in the Ascomycota. Typically, paraphyses are packed between the spore-producing asci. They are often boring: about as long as the asci, skinny, and tubular, with rounded tips. However, in some mushrooms paraphyses are more distinctive, and their anatomy can be a very informative character for mushroom identification. "Hooked" paraphyses (see the illustration to the left) help to define the genus Otidea. Paraphyses in Gyromitra are filled with orangish contents, helping to separate mushrooms in that genus from look-alikes. Paraphyses in Ionomydotis irregularis develop swollen, spear-shaped tips. However, the FPA (Funkiest Paraphyses Award) probably goes to Microstoma floccosum, in which the paraphyses crawl among the asci and encase them. Identification of species of Geoglossum, all of which look more or less the same to the naked eye, relies heavily on the fine points of paraphysis morphology. See also using a microscope. |
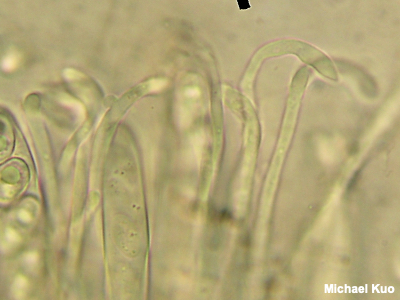
|
|
Parasite, Parasitic Mushrooms that are parasitic consume the living tissues of other organisms (plants, trees, even insects or other mushrooms), sometimes causing a disease or killing them in the process (in which case they are pathogenic). Parasitism is fairly common among mushrooms, and is famously demonstrated in Pseudoboletus parasiticus, which attacks the fungus Scleroderma citrinum. Other parasites include species of Hypomyces, which parasitize mushrooms, and Armillaria solidipes, which attacks trees and is a constant concern in forest management. Species of Cordyceps and related genera win the prize for most astonishing fungal parasites, since they attack insects, as in the illustration to the left. Cordyceps militaris is a well-known example. Readers who have seen the BBC's Planet Earth series have watched Cordyceps mycelium-infected, living ants turned into zombies and commanded by the fungus to climb to an advantageous height before being killed, at which point the mushroom erupts through the dead ant's head to disperse spores into air currents. Compare with mycorrhizal and saprobic. |

|
|
Partial Veil A partial veil is a covering over the gills or pores of young, button mushrooms, serving to protect the spore-bearing surfaces until the spores have matured. The partial veil may take the form of a flimsy tissue which peels away as the mushroom matures, either disappearing or collapsing around the stem to form a ring or ring zone—or it may take the form of a cortina. Independent of whether the partial veil leaves evidence of itself on the mushroom's stem as a ring or ring zone, it may also leave fragments clinging to the margin of the cap (this is often true in Psathyrella candolleana, for example). Sometimes, these partial veil remnants are the only evidence one has that there was ever a partial veil at all, and their presence or absence can be important in mushroom identification. Compare with universal veil. |

|
|
Patches Patches are proportionally large, membranous remainders of a universal veil, left on the cap surface, as in the illustration of Amanita ceciliae to the left. The line separating patches from warts is a bit blurry; the distinction is primarily a matter of size and consistency. Patches tend to be larger and less consistent, while warts are smaller and more consistent. Some mushrooms are known for (usually, or at least often) featuring one large patch (for example Amanita calyptroderma). Compare with warts. See also universal veil. |

|
Pathogen, Pathogenic See parasite. |
|
Pavement Cells See brachybasidioles. |
|
Pendant Hanging like a skirt. See annulus, ring. |
|
|
Perithecium, Perithecia Perithecia (singular: perithecium) are the spore-producing pockets featured in some fungi in the Ascomycota. They are bump- or flask-like structures, often embedded in the surface, or just below the surface, of the mushroom. Inside the perithecium, asci and spores are produced; eventually, when the fungus is mature, the spores are released. Species in the Sordariomycetes feature perithecia; among the most familiar species for mushroom collectors are Xylaria polymorpha and other Xylaria species, Cordyceps militaris, and Daldinia childiae. |

|
|
Peronate A mushroom's stem is "peronate" when it appears booted—like it is wearing a stocking. The stocking is usually the result of a universal veil that initially covered the whole mushroom but, as the stem elongated and the cap expanded, was left on the lower portion of the stem. Species of Cortinarius often have peronate stems, along with some species of Tricholoma, Suillus, Hygrophorus, and other mushrooms. See also annulus, ring and universal veil; compare with volva. |

|
|
Phylogenetic Species, Phylogenetic Species Concept The reigning way of defining a mushroom species at the time of this writing (2018) uses a phylogenetic species concept; species defined this way are phylogenetic species. To make a very long and complicated story short, a phylogenetic species is determined by analysis of its DNA. The science behind DNA analysis of mushrooms is still evolving, and changes fairly dramatically with regularity. Not too long ago (like, only 15–20 years ago), one type of DNA analysis was widely used and papers were published based on these findings; now technological and protocol advances have rendered these findings quaint and often inaccurate. DNA studies looking at one gene were published . . . then two genes became the standard, and now at least three or four are often required (depending on the mushrooms). In short, we can expect further refinements and changes and, with them, revisions of previous findings. If this frustrates you, it may be worth remembering that science would not be science without such progressions and revisions; it would be fundamentalism, instead. Morchella cryptica, pictured to the left, is a well-supported phylogenetic species, but it fails miserably as a morphological species, since its physical features are indistinguishable from those of Morchella esculentoides—which makes it a cryptic species, since you can't tell it from another species (hence its Latin name). Compare with morphological species and biological species. |

|
|
Pileipellis The pileipellis is the surface of a mushroom's cap, as seen under the microscope. There are many types of pileipellis. A cutis is a type of pileipellis in which hyphae are arranged more or less parallel to the cap's surface (illustrated on the page for Pluteus longistriatus); in an ixocutis the hyphae are gelatinized (see Leratiomyces squamosus var. thraustus). A trichoderm, illustrated to the left, is a type of pileipellis in which the hyphae arise perpendicular to the cap surface; in an ixotrichoderm the hyphae are gelatinized (see Hygrocybe glutinipes). In an epithelium the hyphae are also perpendicular to the cap surface, but they are swollen and frequently septate (see Lactarius lignyotus). A hymeniform pileipellis is one in which the hyphae are club-shaped and arise perpendicular to the cap surface; the hyphae are inflated and resemble immature basidia (see Lacrymaria velutina). A pileipellis is sometimes called "cellular" when it appears to consist of swollen cells; a hymeniform pileipellis, for example, can appear "cellular" when viewed from above. In a hyphoepithelium, a trichoderm is covered by a very thin, cutis-like layer. In a lamprotrichoderm the upright elements are thick-walled, elongated, and pointed (see Lactarius subvellereus var. subdistans). See also Using a microscope: the pileipellis. |
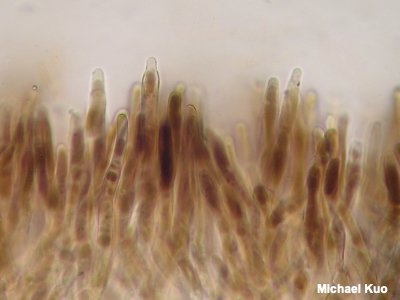
|
Pileocystidia See cystidia. |
|
|
Pileus, Pileate, Cap, Head A pileus (plural: pilei) is the cap, or cap-like structure, of a mushroom; a mushroom that features a cap or head is called "pileate." Mushrooms can feature caps regardless of whether a stem is also present. Caps range from tiny and ephemeral (as in Leucocoprinus fragilissimus, illustrated to the left) to large structures that persist for years (as in Ganoderma applanatum). The Latin suffix —cephala (—cephalo, —cephalus) indicates a mushroom's cap. Thus Lysurus corallocephalus has an coral-like cap. Compare with effused-reflexed and resupinate. |

|
Pleurocystidia See cystidia. |
|
|
Pores, Pore Surface Pores on mushrooms are tiny holes, just as they are with skin. Many mushrooms produce their spores on the inside surfaces of tubes; the mouths of the tubes form pores. The pore surface is the surface comprised of all the tube mouths, together, as in the pore surface of Rubroboletus dupainii, illustrated to the left. Observation of the pore surface is often important in mushroom identification. The size of the pores (usually expressed as a number of pores per millimeter) can be important, as well as their arrangement. Often the tiny holes make no particular pattern, but in some cases they are clearly elongated and radially arranged, especially near the stem. Pores like this, in Mycologese, are called boletinoid (see Boletinelluis merulioides for an example). In some cases pores are so boletinoid that they begin to form ridges near the stem and can approximate the appearance of gills; this kind of arrangement is called lamellate. The color of a mushroom's pore surface can be crucial to identifying it—but be aware that the color is frequently subject to change as the mushroom develops. When identifying boletes, therefore, it is often necessary to have collected mushrooms in both "button" and mature stages. Whether the pore surface bruises is also an important feature; scratch the pore surface with the tip of a knife to see any color changes. The pore surface to the left has bruised blue where it was handled. See also tubes. |

|
Potassium Hydroxide See KOH. |
|
Potholes See scrobiculate. |
|
|
Protologue, Basionym The original, first-ever scientific name for a mushroom species is called the "basionym," and the published text in which the basionym occurs is the "protologue." To the left is the 1753 protologue for Phallus impudicus, which appeared in Linnaeus's Species Plantarum—the first publication to apply what we now call the Linnaean system of binomial nomenclature. Linnaeus gives the species its name ("PHALLUS" and, in the right margin, "impudicus"), then describes it as a Phallus species with a volva, a stem, and a chambered cap. After that he cites five previous descriptions of the mushroom; note that the previous authors did not use the binomial system, with the exception of "Phallus hollandicus f. batavicus (the Batavian regional form of the Dutch Phallus)—and, note the amusing last citation, for "stinky fungus that brings to mind a penis." Then Linnaeus closes the protologue with "Habitat in woods." |

|
|
Pseudostipe, Pseudostem A pseudostem ("pseudostipe" in Mycologese) is, well, pretty much what you would think it is: a not-quite stem. Pseudostems develop on many mushrooms, including some puffballs and species of Scleroderma, some polypores, and some cup fungi. Compare with Stipe, stem, stalk and sterile base. |
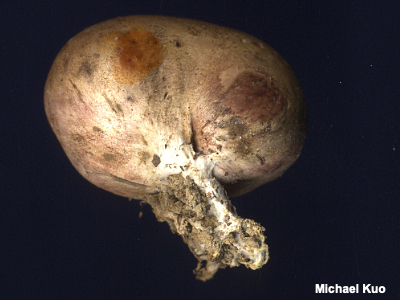
|
|
Resupinate A resupinate mushroom is flattened or closely appressed to the substrate (usually a log), lacking a well defined cap or stem. Resupinate mushrooms are found among crust fungi and polypores—and, depending on what you're calling a mushroom, potentially many other fungi. Many polypores and crusts are opportunistic and develop different fruiting body forms depending on which form (resupinate, effused-reflexed or pileate) will best increase the surface area of the spore-bearing surface. Compare with effused-reflexed and pileate. |
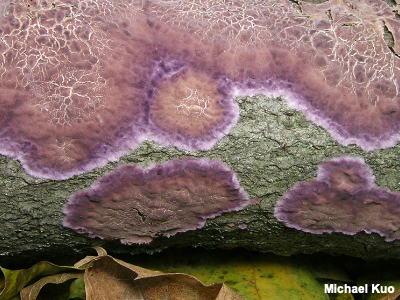
|
|
Reticulation, Reticulate, Reticulated, Reticulum Reticulation is a net-like pattern; something that features reticulation is reticulate, or reticulated. The stems of boletes are often reticulate, and assessing this can be very important in bolete identification. The tiny ridges comprising reticulation on bolete stems are in fact extensions of the pore surface; the criss-cross pattern formed by the reticulation is like the pattern of the pores, but stretched out as a result of the stem's growth (if you're having trouble believing this, see this illustration of the stem of Boletus separans). Although bolete stem reticulation is often obvious, deciding whether some boletes' stems are reticulate, partially reticulate, slightly reticulate near the apex, or not reticulate at all can be one of mushroom identification's more frustrating tasks. Spores of mushrooms sometimes feature reticulate ornamentation; for example this is often the case in Lactarius and Russula, where the reticulation, when present, is amyloid. The word "reticulum" is a noun referring to the network pattern itself—for example, a "stem with a raised reticulum that darkens on handling." |

|
Rhizoids, Rhizomorphs See mycelium. |
|
Ring See annulus, ring and ring zone. |
|
|
Ring Zone A "ring zone" is a zone on the upper stem of a mushroom resulting from the collapse of the partial veil. Ring zones are frequently not as prominent as the rust-colored zone on the Cortinarius species illustrated to the left, which captured rusty spores as the gills matured. Ring zones can be barely noticeable. Use a hand lens if you are unsure! The ring zone may be the result of a tissue-like partial veil, or it may result from a cortina. In my experience, mushrooms that are "supposed to" have ring zones frequently do not, even when a hand lens is used—all the more reason to have both mature and immature specimens available when attempting a difficult identification, since the partial veil will be more easily seen on buttons. Compare with annulus, ring. |

|
|
Saprobe, Saprobic, Saprophyte, Saprotrophic Mushrooms that are saprobes survive by decomposing dead or decaying organic material and using it as food. Many wood-rotting fungi are saprobes, and help decompose deadwood—but other wood rotters are parasitic and attack living wood. Most yard and garden mushrooms (for example Marasmius oreades) are saprobes, as well as dung-loving mushrooms (Panaeolus semiovatus is an example) and mushrooms that decompose leaf or needle litter (like Marasmius pulcherripes). Compare with mycorrhizal and parasite. |

|
|
Scabers, Scabrous Scabers are small, scurfy aggregations of fibers (as on the stem of the Leccinum pictured to the left); something is scabrous if it has scabers. The term comes into play primarily in Leccinum, where the color of the scabers is/was often used as an informative character in identification keys and older taxonomic schemes. Compare with glandular dots. |

|
|
Scrobiculate, Scrobicules, Scrobiculi, Potholes A scrobiculate surface is one that features small pothole-like depressions known as scrobicules or scrobiculi. Generally the term is applied in discussions of the stem surface in Lactarius, where determining the presence or absence of potholes can help in the identification process. However, a Lactarius cap can also feature potholes. Potholes are the result of a thin layer of slime, and represent areas where the cells on the stem surface have become gelatinized. A study of Lactarius stems by Nancy Weber "came to the conclusion that the viscid areas were the smooth shiny scrobiculi present on many species" (Hesler & Smith 1979). Under the microscope, a layer of gelatin-like material can be seen over the potholes. |

|
|
Septate, Septum, Septa A septate structure is divided into sections by cross-walls, which are called septa (singular: septum). Various hyphae can be septate, as well as some spores. In the illustration to the left, the upper spore has six septa, while the bottom spore has seven. In the jelly fungi, it is often useful to know whether or not the basidia are septate or not (and, if so, how) in order to determine the mushroom's genus. Sometimes hyphae become a little swollen and appear constricted at the septa; occasionally this is important information for identification (for example in the paraphyses of Geoglossum species). See also using a microscope. |

|
|
Setae, Seta Setae (singular: seta) are cystidia that are thick-walled and dark brown to black in KOH. The term is usually applied in the polypores (for example in Phellinus and Inonotus) and in the crust fungi (where, for example, "if it ain't got setae, it ain't Hymenochaete"), but some mycologists use the term for similar structures in mushrooms with gills (for example in Parasola conopilus). See also cystidia and using a microscope. |
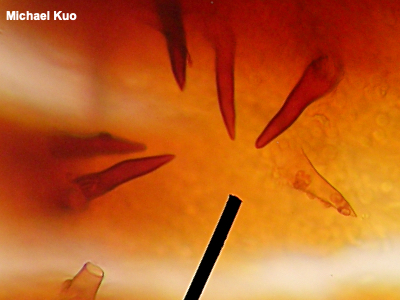
|
|
Short-Gills, Lamellulae Short-gills ("lamellulae" in Mycologese) are gills that begin at the margin of the cap but do not extend all the way to the stem. The presence or absence of short-gills is occasionally an important piece of the mushroom identification puzzle (for example in the genus Russula). Compare with gills and false gills. |

|
Skeletal Hyphae See hyphae. |
|
Skeletocystidia See cystidia. |
|
|
Slime Veil A slime veil is a form of universal veil in which the protective veil is composed of gluten rather than tissue, as in Cortinarius collinitus. Slime veils are found in some species of Cortinarius, Hygrophorus, Gomphidius, and Limacella. In some species of Hygrophorus, like Hygrophorus olivaceoalbus there are two layers to the universal veil: a slime veil and a slightly more tissue-like veil of fibers beneath it. See also universal veil. |

|
|
Spores Spores are microscopic, single-celled units produced by mushrooms in the process of sexual reproduction—roughly analagous to "seeds." The spores are generated on the mushroom's hymenium, and are produced either by asci or basidia, depending on the type of mushroom (officially, spores in the Ascomycota are called "ascospores," and spores in the Basidiomycota are called "basidiospores"). Spores are launched into air currents; eventually, if conditions allow, they germinate to form an organism's new mycelium. Mushroom spores are incredibly diverse, ranging from boring and round or ellipsoid to star-shaped (like the spores of Inocybe insignis to the left), cube-shaped (see Entoloma quadratum), needle-shaped (see Cudonia circinans), spiny (see Strobilomyces confusus), prickly-pear-ish (see Thelephora anthocephala), and so on and so on. Asexual spores, called conidia or, in some cases, chlamydospores, are produced by some mushrooms; conidia can germinate to produce a new organism through cloning, without sexual reproduction. The presence of conidia and conidia-producing structures is sometimes an informative character in mushroom identification (for example in Ganoderma). Illustrations comparing conidia and ascospores of the same species can be found on the page for Kretzschmaria deusta. Assessing the morphology of a mushroom's spores is one of the fundamental elements of mushroom identification; sporal shapes, textures, and measurements are often very crucial to figuring out what a mushroom is. See viewing and measuring spores for an introduction. En masse, spores are visible to the naked eye in the form of a spore print. See also Using a microscope: viewing and measuring spores. |
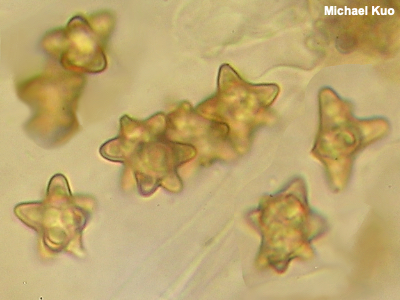
|
|
Spore Print, Spore Deposit A "spore print," sometimes called a "spore deposit," is a mass of spores that can be seen with the naked eye. The white spore print of Armillaria mellea can be seen in the photo to the left; because these mushrooms grow in clusters, they often overlap, so that the gills sit above the surfaces of other caps during development, resulting in a spore print if the gills reach maturity. The color of a mushroom's spore print is essential information in the identification process. Fortunately it is possible to make a spore print, rather than rely on nature to produce in-situ prints like the ones illustrated. See making spore prints for an introduction to the process. For centuries mushrooms were classified in great part according to the color of their spore prints. While recent DNA-based mycology has shown that such arrangements were not necessarily always accurate (for example, the white-spored Lepiota cristata is more closely related to the black-spored Coprinus comatus than it is to many other white-spored mushrooms), spore print color is still one of the best tools in the identification toolbox, regardless of whether the colors represent natural groupings. See also spores and making spore prints. |

|
|
Spore Case A spore case is pretty much what it sounds like: a casing that holds spores. The term is generally used for puffballs. Usually the spore case is initially filled with flesh-like material; with maturity spores are produced and the fleshy consistency gives way to a powdery mass of spores. The spores are released into air currents through an opening or perforation that may develop in the spore case—or the spore case may basically disintegrate, partially or altogether. |

|
|
Sterile Base A sterile base is the basal, fleshy portion of the interior of some puffballs, separate from the spore-producing gleba. The presence or absence of a sterile base can be a useful character in identifying puffballs. Species of Lycoperdon, for example, usually feature a fairly well-developed sterile base, while species of Bovista do not. Compare with gleba, stem, and pseudostipe. |
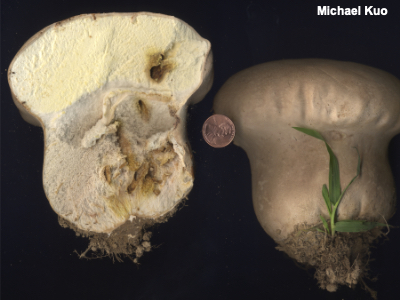
|
|
Stipe, Stipitate, Stem, Stalk In Mycologese, the "stipe" is the stem, or stalk, of a mushroom; a mushroom with a stem is "stipitate." Stems are incredibly variable among mushrooms—and in many mushrooms the stem changes rather dramatically in the course of a mushroom's development. the morphology of a mushroom's stem is often crucial in the identification process: the shape, dimensions, colors, and surface texture can all be important. When collecting mushrooms with small, fragile stems for identification, be careful not to handle the stems; fine details (an ephemeral ring zone, for example) can be accidentally obliterated with a slip of a finger. The genus Psathyrella is known in part for its fragile, easily-snapped-in-two stems; my friend Bob Zordani calls them "snapyrellas." The Latin suffix —ipes indicates a mushroom's stem. Thus Retiboletus ornatipes has an ornate stem. The Latin prefis Caulo— also refers to the stem; "caulocystidia," for example, are cystidia on the stem surface. Compare with pseudostipe. |

|
|
Teleomorph, Teleomorphic The life cycle of some fungi involves both sexual and asexual stages. For such fungi the teleomorph is the sexual stage, while the anamorph is the asexual stage. In the teleomorphic stage sexual reproduction occurs through the production of sexual spores. In older taxonomic schemes anamorphs and teleomorphs were sometimes placed in different genera, despite being life cycle stages of the same organism. For example, the anamorph to the left was often recognized as "Xylocoremium flabelliforme," while its teleomorph was Xylaria cubensis. However, more current taxonomic rules disallow this practice, and the name Xylocoremium flabelliforme is now a deprecated synonym for Xylaria cubensis. Compare with anaomorph. |

|
Trichoderm See pileipellis. |
|
Trimitic Hyphal System See hyphae. |
|
|
Tubes To increase the area of the spore-producing surface some mushrooms have developed the ingenious strategy of using tubes. Imagine covering the inside surface of a paper towel roll (the cardboard cylinder, once the roll is empty) with seeds—then affixing many such tubes to a flat surface. Now compare the number of seeds you would use to the number required to simply cover the flat surface itself. Many, many more seeds are involved with the tubes. Tube walls are usually fused lengthwise, but in Fistulina hepatica and Pseudofistulina radicata they are not fused and can be observed clearly as individual, discrete tubes. "Normal" bolete tubes are like the tubes pictured to the left and, when teased apart, they separate into packed sections. Another tube type is xerocomoid tubes. Tubes are used by boletes and polypores, as well as a few other mushrooms, to hold the spore-producing basidia. At maturity the basidia release the spores, which fall out through the mouths of the tubes (the pores) and into air currents. (In the photo to the left the mushroom is turned upside down, so that the cap is on the bottom.) Some polypores are long-lived and produce new tube layers annually. See the page for Ganoderma applanatum for an illustration. See also hymenium, xerocomoid tubes, and pores, pore surface. |
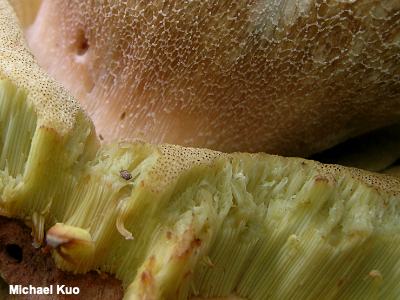
|
|
Type Collection, Type Species A type collection is the mushroom collection designated by the person who names a species as the representative collection for the species. Type collections are deposited in research collections of public museums or herbaria so that future scientists can refer to the original concept of a species. The type collection of Morchella prava is pictured to the left; it is now deposited in the herbarium of the Field Museum of Natural History, in Chicago. Strictly speaking, what I am calling a "type collection" is called the holotype collection, or just the holotype. There are other kinds of type collections: an isotype is a duplicate of the holotype (for example a collection made in the exact same location as the holotype); a paratype is an additional collection cited by the original author when she named the species; and epitypes, neotypes, and lectotypes are various type collections used to correct the problem of a missing, problematic, or misunderstood holotype. A type species is the species chosen by a mycologist to represent a genus. For example, Boletus edulis is the type species of the genus Boletus—which is why only it and very similar, closely related species will retain the genus name Boletus as DNA studies sort out the rest of the former species of "Boletus," which, it turns out, are not so closely related to Boletus edulis. |

|
|
Universal Veil A universal veil is a protective layer of tissue that completely encloses a developing mushroom; the universal veil of Coprinopsis variegata is illustrated to the left: the universal veil is just beginning to rupture on the closest specimen, and the grayish material underneath is the developing mushroom's cap, inside the veil. The rupturing universal veil may disappear as the mushroom expands and matures, or it may result in warts or patches on the cap, and/or a volva at the stem base. A slime veil is a universal veil composed of gluten rather than tissue. An appendiculate cap has universal veil tissue hanging from its margin. Many mushrooms display a universal veil, including Amanita species like Amanita muscaria, Volvariella species, and stinkhorns. Developing mushrooms that are completely encased in a universal veil are sometimes mistaken for puffballs. However, slicing the mushroom open will quickly clear up confusion: while a developing mushroom can be easily seen in cross-section, a puffball's interior is either homogenous and fleshy, or there are two fleshy areas (the gleba and the sterile base). Compare with partial veil. See also appendiculate, patches, slime veil, volva, and warts. |

|
|
Viscid Viscid is Mycologese for "slimy" or "sticky." Officially, a viscid surface is thinly slimy, while a thickly slimy surface is glutinous, but the distinction is iffy and often ignored. A mushroom's surfaces—especially the cap and stem—can be slimy when the specimen is fresh, and the sliminess can be important in mushroom identification. The problem is, mushrooms frequently dry out quickly, making it hard to tell whether it was once slimy. One clue involves appressed, dried out forest litter stuck to the cap surface. In the illustration to the left, leaf fragments are embedded in the gluten that makes the fresh cap of Hygrophorus paludosus slimy. Imagine what would happen if the cap were to dry out over the course of a few days: the leaf fragments would become stuck to the surface, even though the gluten has disappeared. Under the microscope it is also possible to check the "slime factor" after a mushroom has dried out: slimy caps usually correspond with an ixocutis or ixotrichoderm. Assessing the slime factor can be important in many kinds of mushrooms, but it is often involved with identification of russulas and species of Cortinarius and Suillus. |

|
|
Volva, Volvate A volva is the remainder of a universal veil at the base of a mushroom's stem; the volva results from the growing mushroom pushing through the veil. A mushroom with a volva is "volvate." Volvas are found in Amanita, Volvariella and Volvopluteus, and the stinkhorns—and occasionally elsewhere. Volval anatomy is often crucial when identifying amanitas. The "classic" volva is prominent and sack-like, as in Amanita jacksonii or Amanita phalloides. However, volvas are diverse. In Amanita muscaria var. flavivolvata the volva consists of prominent concentric ringlets of tissue left at the top of the stem's basal bulb. Volvas can form a little roll of tissue at the top of the stem's bulb, in which case they are called limbate (the roll of tissue is a limb); see Amanita multisquamosa for an example. In some species the volva is friable and breaks up into fragments of tissue that adhere to the stem base (or fall off onto the ground around the mushroom), as in Amanita flavoconia. And in some species the volva is barely discernible, left as a mere smear on the stem base, as in Amanita farinosa. Compare with partial veil. See also universal veil. |

|
|
Warts Warts are small, membranous remainders of a universal veil, left on the cap surface after the mushroom's growth has ruptured the veil and stretched it out. Warts are similar to patches, but they are smaller and more consistently arranged. Warts can be washed off with rain, making them a difficult feature to be sure of. Warts are found in the genus Amanita, and occasionally elsewhere. Compare with patches. See also universal veil. |

|
|
White Rot The mycelia of white rot fungi degrade lignin in the wood they inhabit, but not the cellulose, resulting in a characteristic whitish, stringy decay of the wood. White rot fungi are usually found on the wood of hardwoods. Well-known white rot fungi include species of Armillaria, Pleurotus ostreatus, and Ganoderma applanatum. Compare with brown rot. |

|
|
Xerocomoid Tubes Some boletes have xerocomoid tubes—meaning the tubes are not easily separable; if you try to tease them apart, they rip, rather than separating more or less easily into sections of packed-together tubes. Xerocomoid tubes are primarily (but not exclusively) found in Xerocomus and Xerocomellus. See also tubes. |

|
|
Zonate A zonate surface features concentric zones of color and/or texture, as in the cap of Lactarius indigo, illustrated to the left. Usually it is the cap that is described as zonate, but sometimes mushrooms featured zonate flesh, as well (see for example Fomitopsis spraguei). |

{kind=link}
|
This site contains no information about the edibility or toxicity of mushrooms. Note: To provide a link to a specific entry on this page, use the format www.mushroomexpert.com/glossary.html#__________ and fill in the blank with the entry's term/label, using lower case letters and eliminating spaces or using an underscore ( _ ) in their place. Kuo, Michael (2019). Glossary. Retrieved from the Mushroomexpert.Com website: www.mushroomexpert.com/glossary.html All text and images © , Mushroomexpert.com |